Сборка эндоплазматического ретикулума
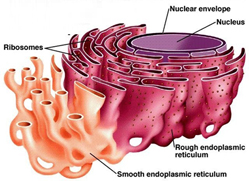
На рисунке — эндоплазматический ретикулум (ER), черные точки — рибосомы.
У ER много функций, основные из которых: синтез белков, стерола, липидов; накопление и регулируемое высвобождение Ca 2+; детоксикация.
Соотношение гладкого и шероховатого доменов ER
Это соотношение может меняться как динамически, в режиме реального времени, так и стабильно отличаться в разных типах клеток. В частности, площадь гладкого ER начинает увеличиваться в присутствии токсинов, таких как этанол. Это повышает синтез цитохрома P450 и других детоксифицирующих ферментов.
В свою очередь, грубый ER расширяется, когда секреция белка увеличивается. Например, во время дифференцировки B-лимфоцитов в плазматические клетки.
В целом, все секреторные клетки обнаруживают большие объемы шероховатых листов. Напротив, гепатоциты печени характеризуются обширной сетью гладкого ретикулума, которая обеспечивает метаболизм и детоксикацию.
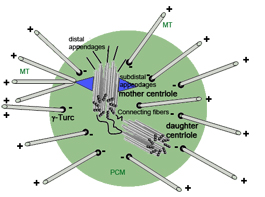
Взаимодействие с микротрубочками
На рисунке – клеточное ядро. Складывается впечатление, что оно пронизано полярными микротрубочками. На самом деле это не так: микротрубочки связываются с белками на поверхности ядра.
Эта тенденция верна и для эндоплазматического ретикулума: микротрубочки связаны с поверхностью его листов и не проникают внутрь.
Домены ER
Эндоплазматический ретикулум не является органеллой с непрерывной мембраной, а состоит из отдельных мембранных доменов. В первую очередь это гладкий и шероховатый ретикулум, а также многочисленные внешние домены, формируемые на базе гладкого ER [1].
Существуют различные доказательства, что полости гладкого и шероховатого ретикулумов не сообщаются. Так, фермент триацилглицерингидролаза возвращается в гладкий ретикулум из Гольджи. При этом стандартной сигнальной последовательности ретикулума — KDEL — ей недостаточно, ей нужна другая адресная последовательность: гистидин-изолейцин-аспарагиновая кислота-лейцин (HIEL).
Все это хорошо вписывается в наблюдаемое различие белков в разных доменах. Но что это означает? Отдельные полости как у Гольджи? Или пограничный контроль на стыках мембран?
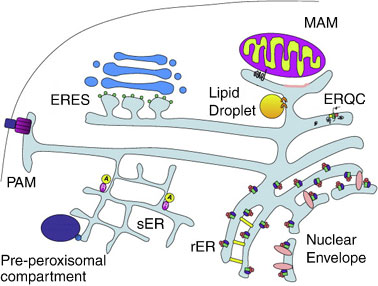
Общая схема сборки ER
Гладкий ER существует всегда. В телофазе на его базе собираются мембрана ядерной оболочки и шероховатый ER.
Далее ER генерирует de novo 3 вида органелл: комплекс Гольджи, липидные капли и пероксисомы. Под эти задачи на гладком ER формируются соответствующие домены.
Также на базе гладкого ER формируются еще 3 домена:
— митохондриально-ассоциированная мембрана (MAM);
— компартмент контроля качества ретикулума (ERQC);
— мембрана, ассоциированная с плазматической мембраной (PAM).
1. Гладкий ER
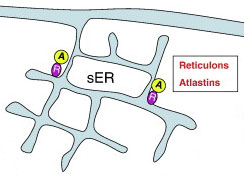
Как отмечалось, гладкий ER существует всегда. На его базе формируются почти все остальные домены ER. Гладкий ER реализует как минимум 2 функции:
1. Синтез липидов.
2. Хранение кальция. Обеспечивается кальцийсвязывающими белками кальретикулинами, кальциевыми насосами SERCA, рецепторами IP3R.
На рисунке: гладкая эндоплазматическая сеть (sER), ретикулоны (фиолетовые круги) и атластины (желтые круги).
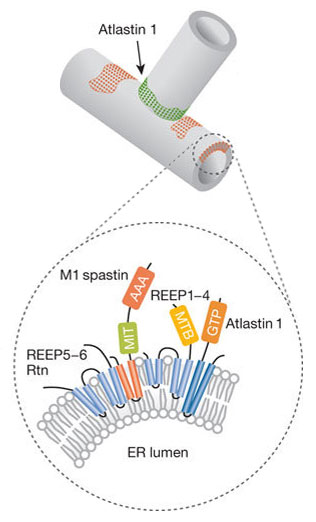
Существует несколько классов белков, которые являются необходимыми и достаточными для образования и поддержания ER канальцев в эукариотических клетках. А именно:
— атластины — RHD3 / Sey1, для формирования трехсторонних соединений;
— ретикулоны;
— DP1 / REEPs / Yop1, несущие гидрофобные шпилечные домены.
Равновесие между атластинами, ретикулонами и DP1 / Yop1p определяет образование гладких канальцев ER [2].
Представленные на рисунке белки spastin и REEP1-4 обеспечивают взаимодействие канальцев ER с цитоскелетом микротрубочек.
2. Сборка мембраны ядерной оболочки
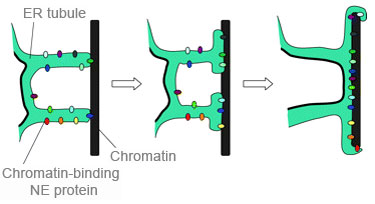
1. У млекопитающих в ранней телофазе трубчатые выступы гладкого ER связываются с хроматином, инициируя процесс сборки ядерной NE-оболочки [3].
Это возможно, т.к. многие ER-ассоциированные белки внутренней ядерной мембраны обладают высоким сродством к ДНК.
Точно не известно, какие именно белки ER первыми свяжутся с хроматином. Но известно, что с хроматином умеют связываться, например, LBR и Lap2B. Причем, LBR умеет связываться непосредственно с ДНК.
2. После этого начального связывания мембранные канальцы сплющиваются в листы, которые распространяются по хроматину и реорганизуются в герметичную NE-оболочку.
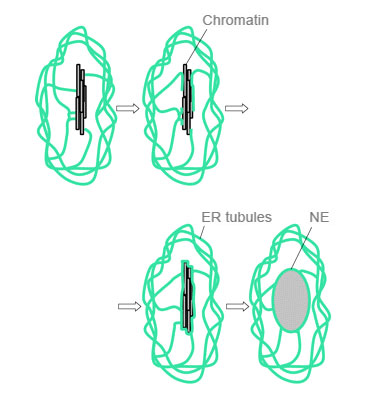
На рисунке — то же самое, в более крупном масштабе.
3. В единое ядро объединяются только хромосомы связанные с веретеном деления. Хромосомы, не связавшиеся с веретеном, в телофазе организуют свои микроядра, которые уже не смогут слиться с основным ядром.
Как гладкий ER родителя делится между двумя дочерними клетками?
Находится ли ядро под каким-нибудь давлением? Или сама распакованная ДНК придает ядру форму по типу «оболочка наполненная ватой»?
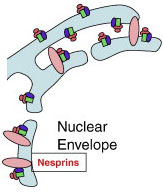
На рисунке – переход от ядерной оболочки к шероховатому ER.
И внутренняя и внешняя ядерные мембраны отличаются от мембран шероховатого ER уникальным набором белков.
На внешней ядерной мембране размещены рибосомы и белки семейства Nesprins (розовые овалы). Несприны соединяют ядерную оболочку с цитоскелетом. Также они взаимодействуют с белками внутренней ядерной мембраны используя домен KASH (klarsicht / ANC-1 / syne).
3. Сборка шероховатого ER
Экспансия шероховатого ER всегда начинается с синтеза листов на базе гладкого ER, которые позже могут быть украшены рибосомами.
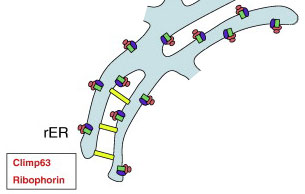
Листы шероховатого ER (rER) содержат рибосомы (розовые овалы), которые связаны с компонентами транслокона (зеленые прямоугольники) и рибофоринами (рибофорин I / II, фиолетовые овалы).
Мембранный белок Climp63 (желтый прямоугольник) является важным структурным белком rER. Он формирует плоские листы ER, к которым могут прикрепляться рибосомы. Постоянная толщина просвета листов ER стабилизируется внутрипросветными мостиками, образованными Climp63.
Еще один белок, p180 играет центральную роль в прикреплении рибосом к ER-листам в клетках млекопитающих. И Climp63, и p180 связывают ER с микротрубочками цитоскелета.
Также с шероховатым ER ассоциируются белки Sec61 и Sec63.
4. Домен ERES для взаимодействия с Гольджи
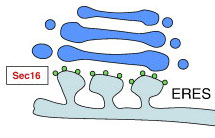
ERES — это основной порт секреторного пути. Здесь покрытые COPII везикулы отрываются от ER и движутся в сторону Гольджи.
На полпути к Гольджи ERES генерирует промежуточный компартмент — ER-Golgi (ERGIC).
В формировании везикул участвуют белки Sec16A, Sec16B (маленькие зеленые кружки). Подробнее об этом см. формирование везикул в рубрике «Путь белка».
5. Митохондриально-ассоциированная мембрана (MAM)
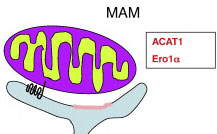
MAM (Mitochondria Associated Membrane) представляет собой участок гладкого ER, который функционирует как порт для работы с митохондриями. На сегодня известно, что до 20% поверхности митохондрий находится в контакте с ER.
MAM обеспечивает обмен липидов между ER и митохондриями. Соответственно на нем можно обнаружить большое количество липидсинтезирующих ферментов, а также микродомены, обогащенные гликосфинголипидами.
Также MAM выполняет центральную роль в высвобождении кальция из ER.
На рисунке: белок ACAT1 (черная волнистая линия) и другие белки (розовый участок мембраны). С MAM ассоциируются такие белки как UL37, DGAT2, BAP31, FACL4, Erolα, Rab32, Mfn2, VDAC, PACS2, Mmm1, Mdm10, шапероны Grp75, рецепторы IP 3 и Sigma-1.
6. Компартмент контроля качества ретикулума (ERQC)
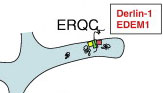
В ERQC происходит экспорт развернутых белков в протеасому – «ER-ассоциированная деградация белков» (ERAD).
ERQC является производным от шероховатого rER. Этот домен сохранил компоненты транслокона под рибосомы (зеленый прямоугольник), хотя использует их в противоположном направлении. Также содержит связанные с транслоконом белки Derlin-1 (желтый квадрат) и EDEM-1 (розовый квадрат), которые облегчают убиквитинирование и ретротранслокацию неправильно свернутых белков из ER (подробнее см. в рубрике «Прочие процессы»).
Шаперон калнексин не входит непосредственно в состав ERQC. Но при стрессах вида «тепловой шок» может сюда переместиться из MAM.
В процессах участвуют BAP31.
7. Мембрана, ассоциированная с плазматической мембраной (PAM)
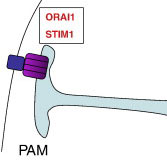
PAM широко взаимодействует с плазматической мембраной в целых 1100 контактных сайтах и реализует несколько функций:
1. Обеспечивает импорт кальция из внеклеточного пространства. Для этого STIM1 (пурпурный прямоугольник) олигомеризуется с образованием поры, которая ассоциируется с кальциевым каналом ORAI1 плазматической мембраны (темно-синий прямоугольник).
2. Транспортировка стеролов из ER к плазматической мембране.
3. В клетках дрожжей и млекопитающих PAM является основным местом синтеза фосфатидилсерина. Этим липидом маркируется внутренняя (цитоплазматическая) часть клеточной мембраны.
Фосфатидилсерин играет важную роль во время апоптоза, когда он попадает на внешний листок плазматической мембраны. Это в свою очередь позволяет фагоцитам распознавать апоптотические клетки и поглощать их.
8. Домен биогенеза пероксисом
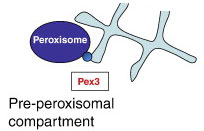
Этот домен синтезирует пероксисомы.
Белки, предназначенные для пероксисом, сортируются в препероксисомальный компартмент, где они отпочковываются в пре-пероксисомальные пузырьки зависимым от Pex3 (маленький синий кружок) образом. Также с данным процессом ассоциируются Pex16 и Pex19.
9. Домен биогенеза липидных капель
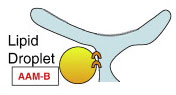
Домен обеспечивает хранение триацилглицеридов.
DGAT1 и DGAT2 катализируют заключительную стадию синтеза триацилглицеридов, но только DGAT2 может перемещаться из МАМ в места биогенеза липидных капель. На этих участках ферментативная активность DGAT2 направляет триацилглицериды в образующиеся цитозольные липидные капли.
Географически липидные капли находятся в тесном контакте с ERES и ERGIC. Готовые липидные капли покрываются белками перилипина.
Кроме перечисленных, с синтезом липидных капель ассоциируются такие белки как AAM-B, PAT, TIP47, PKA.
10. Тельца Рассела
ER может генерировать тельца Рассела, которые хранят мутантные, нерастворимые иммуноглобулины, которые не могут ни секретироваться, ни расщепляться. На начальном этапе формирования телец Рассела иммуноглобулины распознаются ERES-ассоциированными шаперонами ERGIC-53 и ERp44.
Тельца Рассела формируются либо на шероховатом, либо на гладком ER, указывая тем самым, что оба домена приспособлены для удаления нежелательных отходов.
11. Домен GERL
Домен GERL (Golgi-ER-lysosome) формирует особые типы лизосом непосредственно из ER.